Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала.
Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл . Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G 1 , синтетического — S, постсинтетического, или премитотического, — G 2 .
Пресинтетический период (2n 2c , где n — число хромосом, с — число молекул ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду.
Синтетический период (2n 4c ) — репликация ДНК.
Постсинтетический период (2n 4c ) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
Профаза (2n 4c ) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом.
Метафаза (2n 4c ) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4n 4c ) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.
1 — профаза; 2 — метафаза; 3 — анафаза; 4 — телофаза.
Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др.
— это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное. Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК.
Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2n 4c ) образуются две гаплоидные (1n 2c ).
Интерфаза 1 (в начале — 2n 2c , в конце — 2n 4c ) — синтез и накопление веществ и энергии, необходимых для осуществления обоих делений, увеличение размеров клетки и числа органоидов, удвоение центриолей, репликация ДНК, которая завершается в профазе 1.
Профаза 1 (2n 4c ) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом, конъюгация гомологичных хромосом и кроссинговер. Конъюгация — процесс сближения и переплетения гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом . Кроссинговер — процесс обмена гомологичными участками между гомологичными хромосомами.
Профаза 1 подразделяется на стадии: лептотена (завершение репликации ДНК), зиготена (конъюгация гомологичных хромосом, образование бивалентов), пахитена (кроссинговер, перекомбинация генов), диплотена (выявление хиазм, 1 блок овогенеза у человека), диакинез (терминализация хиазм).

1 — лептотена; 2 — зиготена; 3 — пахитена; 4 — диплотена; 5 — диакинез; 6 — метафаза 1; 7 — анафаза 1; 8 — телофаза 1;
9 — профаза 2; 10 — метафаза 2; 11 — анафаза 2; 12 — телофаза 2.
Метафаза 1 (2n 4c ) — выстраивание бивалентов в экваториальной плоскости клетки, прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза 1 (2n 4c ) — случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая — к другому), перекомбинация хромосом.
Телофаза 1 (1n 2c в каждой клетке) — образование ядерных мембран вокруг групп двухроматидных хромосом, деление цитоплазмы. У многих растений клетка из анафазы 1 сразу же переходит в профазу 2.
Второе мейотическое деление (мейоз 2) называется эквационным .
Интерфаза 2 , или интеркинез (1n 2c ), представляет собой короткий перерыв между первым и вторым мейотическими делениями, во время которого не происходит репликация ДНК. Характерна для животных клеток.
Профаза 2 (1n 2c ) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления.
Метафаза 2 (1n 2c ) — выстраивание двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом; 2 блок овогенеза у человека.
Анафаза 2 (2n 2с ) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами), перекомбинация хромосом.
Телофаза 2 (1n 1c в каждой клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток.
Биологическое значение мейоза. Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. Являясь основой комбинативной изменчивости, мейоз обеспечивает генетическое разнообразие гамет.
Амитоз
Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл.
Клеточный цикл
Клеточный цикл — жизнь клетки от момента ее появления до деления или смерти. Обязательным компонентом клеточного цикла является митотический цикл, который включает в себя период подготовки к делению и собственно митоз. Кроме этого, в жизненном цикле имеются периоды покоя, во время которых клетка выполняет свойственные ей функции и избирает дальнейшую судьбу: гибель или возврат в митотический цикл.
Перейти к лекции №12 «Фотосинтез. Хемосинтез»
Перейти к лекции №14 «Размножение организмов»
Сопровождающееся уменьшением числа хромосом вдвое. Он состоит из двух последовательно идущих делений, имеющих те же фазы, что и митоз. Однако, как показано в таблице «Сравнение митоза и мейоза» , продолжительность отдельных фаз и происходящие в них процессы значительно отличаются от процессов, происходящих при митозе.
Эти отличия в основном состоят в следующем.
В мейозе профаза I более продолжительна. В ней происходит конъюгация (соединение гомологичных хромосом) и обмен генетической информацией . В анафазе I центромеры , скрепляющие хроматиды, не делятся , а к полюсам отходит одна из гомологмейоза митоза и ичных хромосом. Интерфаза перед вторым делением очень короткая , в ней ДНК не синтезируется . Клетки (галиты ), образующиеся в результате двух мейотических делений, содержат гаплоидный (одинарный) набор хромосом. Диплоидность восстанавливается при слиянии двух клеток - материнской и отцовской. Оплодотворенную яйцеклетку называют зиготой .
Митоз и его фазы
Митоз, или непрямое деление , наиболее широко распространен в природе. Митоз лежит в основе деления всех неполовых клеток (эпителиальных, мышечных, нервных, костных и др.). Митоз состоит из четырех последовательных фаз (см. далее таблицу). Благодаря митозу обеспечивается равномерное распределение генетической информации родительской клетки между дочерними. Период жизни клетки между двумя митозами называют интерфазой . Она в десятки раз продолжительнее митоза. В ней совершается ряд очень важных процессов, предшествующих делению клетки: синтезируются молекулы АТФ и белков , удваивается каждая хромосома, образуя две сестринские хроматиды , скрепленные общей центромерой , увеличивается число основных органоидов цитоплазмы.
В профазе спиралируются и вследствие этого утолщаются хромосомы , состоящие из двух сестринских хроматид, удерживаемых вместе центромерой. К концу профазы ядерная мембрана и ядрышки исчезают и хромосомы рассредоточиваются по всей клетке, центриоли отходят к полюсам и образуют веретено деления . В метафазе происходит дальнейшая спирализация хромосом. В эту фазу они наиболее хорошо видны. Их центромеры располагаются по экватору. К ним прикрепляются нити веретена деления.
В анафазе центромеры делятся, сестринские хроматиды отделяются друг от друга и за счет сокращения нитей веретена отходят к противоположным полюсам клетки.
В телофазе цитоплазма делится, хромосомы раскручиваются, вновь образуются ядрышки и ядерные мембраны. В животных клетках цитоплазма перешнуровывается, в растительных - в центре материнской клетки образуется перегородка. Так из одной исходной клетки (материнской) образуются две новые дочерние.
Таблица - Сравнение митоза и мейоза
| Фаза | Митоз | Мейоз | |
| 1 деление | 2 деление | ||
| Интерфаза |
Набор хромосом 2n. Идет интенсивный синтез белков, АТФ и других органических веществ. Удваиваются хромосомы, каждая оказывается состоящей из двух сестринских хроматид, скрепленных общей центромерой. |
Набор хромосом 2n Наблюдаются те же процессы, что и в митозе, но более продолжительна, особенно при образовании яйцеклеток. | Набор хромосом гаплоидный (n). Синтез органических веществ отсутствует. |
| Профаза | Непродолжительна, происходит спирализация хромосом, исчезают ядерная оболочка, ядрышко, образуется веретено деления. | Более длительна. В начале фазы те же процессы, что и в митозе. Кроме того, происходит конъюгация хромосом, при которой гомологичные хромосомы сближаются по всей длине и скручиваются. При этом может происходить обмен генетической информацией (перекрест хромосом) - кроссинговер . Затем хромосомы расходятся. | Короткая; те же процессы, что и в митозе, но при n хромосом. |
| Метафаза | Происходит дальнейшая спирализация хромосом, их центромеры располагаются по экватору. | Происходят процессы, аналогичные тем, что и в митозе. | |
| Анафаза | Центромеры, скрепляющие сестринские хроматиды, делятся, каждая из них становится новой хромосомой и отходит к противоположным полюсам. | Центромеры не делятся. К противоположным полюсам отходит одна из гомологичных хромосом, состоящая из двух хроматид, скрепленных общей центромерой. | Происходит то же, что и в митозе, но при n хромосом. |
| Телофаза | Делится цитоплазма, образуются две дочерние клетки, каждая с диплоидным набором хромосом. Исчезает веретено деления, формируются ядрышки. | Длится недолго Гомологичные хромосомы попадают в разные клетки с гаплоидным набором хромосом. Цитоплазма делится не всегда. | Делится цитоплазма. После двух мейотических делений образуется 4 клетки с гаплоидным набором хромосом. |
Таблица сравнения митоза и мейоза.
Интерфаза – это период между двумя клеточными делениями. В интерфазе ядро компактное, не имеет выраженной структуры, хорошо видны ядрышки. Совокупность интерфазных хромосом представляет собой хроматин. В состав хроматина входят: ДНК, белки и РНК в соотношении 1: 1,3: 0,2, а также неорганические ионы. Структура хроматина изменчива и зависит от состояния клетки.
Хромосомы в интерфазе не видны, поэтому их изучение ведется электронно-микроскопическими и биохимическими методами. Интерфаза включает три стадии: пресинтетическую (G1), синтетическую (S) и постсинтетическую (G2). Символ G представляет собой сокращение от англ. gap – интервал; символ S – сокращение от англ. synthesis – синтез. Рассмотрим эти стадии подробнее.
Пресинтетическая стадия (G1). В основе каждой хромосомы лежит одна двуспиральная молекула ДНК. Количество ДНК в клетке на пресинтетической стадии обозначается символом 2с (от англ. content – содержание). Клетка активно растет и нормально функционирует.
Синтетическая стадия (S). Происходит самоудвоение, или репликация ДНК. При этом одни участки хромосом удваиваются раньше, а другие – позже, то есть репликация ДНК протекает асинхронно. Параллельно происходит удвоение центриолей (если они имеются).
Постсинтетическая стадия (G2). Завершается репликация ДНК. В состав каждой хромосомы входит две двойных молекулы ДНК, которые являются точной копией исходной молекулы ДНК. Количество ДНК в клетке на постсинтетической стадии обозначается символом 4с. Синтезируются вещества, необходимые для деления клетки. В конце интерфазы процессы синтеза прекращаются.
Процесс митоза
Профаза – первая фаза митоза. Хромосомы спирализуются и становятся видны в световой микроскоп в виде тонких нитей. Центриоли (если они имеются) расходятся к полюсам клетки. В конце профазы ядрышки исчезают, ядерная оболочка разрушается, и хромосомы выходят в цитоплазму.
В профазе увеличивается объем ядра, и вследствие спирализации хроматина формируются хромосомы. К концу профазы видно, что каждая хромосома состоит из двух хроматид. Постепенно растворяются ядрышки и ядерная оболочка, и хромосомы оказываются беспорядочно расположенными в цитоплазме клетки. Центриоли расходятся к полюсам клетки. Формируется ахроматиновое веретено деления, часть нитей которого идет от полюса к полюсу, а часть - прикрепляется к центромерам хромосом. Содержание генетического материала в клетке остается неизменным (2n2хр).
Рис. 1. Схема митоза в клетках корешка лука

Рис. 2. Схема митоза в клетках корешка лука: 1- интерфаза; 2,3 - профаза; 4 - метафаза; 5,6 - анафаза; 7,8 - телофаза; 9 - образование двух клеток

Рис. 3. Митоз в клетках кончика корешка лука: а - интерфаза; б - профаза; в - метафаза; г - анафаза; л, е - ранняя и поздняя телофазы
Метафаза. Начало этой фазы называется прометафаза. В прометафазе хромосомы располагаются в цитоплазме довольно беспорядочно. Формируется митотический аппарат, в состав которого входит веретено деления и центриоли или иные центры организации микротрубочек. При наличии центриолей митотический аппарат называется астральным (у многоклеточных животных), а при их отсутствии – анастральным (у высших растений). Веретено деления (ахроматиновое веретено) – это система тубулиновых микротрубочек в делящейся клетке, обеспечивающая расхождение хромосом. В состав веретена деления входят два типа нитей: полюсные (опорные) и хромосомальные (тянущие).
После формирования митотического аппарата хромосомы начинают перемещаться в экваториальную плоскость клетки; это движение хромосом называется метакинез.
В метафазе хромосомы максимально спирализованы. Центромеры хромосом располагаются в экваториальной плоскости клетки независимо друг от друга. Полюсные нити веретена деления тянутся от полюсов клетки к хромосомам, а хромосомальные – от центромер (кинетохоров) – к полюсам. Совокупность хромосом в экваториальной плоскости клетки образует метафазную пластинку.
Анафаза. Происходит разделение хромосом на хроматиды. С этого момента каждая хроматида становится самостоятельной однохроматидной хромосомой, в основе которой лежит одна молекула ДНК. Однохроматидные хромосомы в составе анафазных групп расходятся к полюсам клетки. При расхождении хромосом хромосомальные микротрубочки укорачиваются, а полюсные – удлиняются. При этом полюсные и хромосомальные нити скользят вдоль друг друга.
Телофаза. Веретено деления разрушается. Хромосомы у полюсов клетки деспирализуются, вокруг них формируются ядерные оболочки. В клетке образуются два ядра, генетически идентичные исходному ядру. Содержание ДНК в дочерних ядрах становится равным 2c.
Цитокинез. В цитокинезе происходит разделение цитоплазмы и формирование мембран дочерних клеток. У животных цитокинез происходит путем перешнуровывания клетки. У растений цитокинез происходит иначе: в экваториальной плоскости образуются пузырьки, которые сливаются с образованием двух параллельных мембран.
На этом митоз завершается, и наступает очередная интерфаза.
Различают следующие четыре фазы митоза: профаза, метафаза, анафаза и телофаза
. В профазе
хорошо видны центриоли
- образования, находящиеся в клеточном центре и играющие роль в делении дочерних хромосом животных. (Напомним, что у высших растений нет центриолей в клеточном центре, который организует деление хромосом). Мы же рассмотрим митоз на примере животной клетки, поскольку присутствие центриоли делает процесс деления хромосом более наглядным. Центриоли делятся и расходятся к разным полюсам клетки. От центриолей протягиваются микротрубочки, образующие нити веретена деления, которое регулирует расхождение хромосом к полюсам делящейся клетки.
В конце профазы ядерная оболочка распадается, ядрышко постепенно исчезает, хромосомы спирализуются и в результате этого укорачиваются и утолщаются, и их уже можно наблюдать в световой микроскоп. Еще лучше они видны на следующей стадии митоза - метафазе
.
В метафазе хромосомы располагаются в экваториальной плоскости клетки. При этом хорошо видно, что каждая хромосома, состоящая из двух хроматид, имеет перетяжку - центромеру
. Хромосомы своими центромерами прикрепляются у нити веретена деления. После деления центромеры каждая хроматида становится самостоятельной дочерней хромосомой.
Затем наступает следующая стадия митоза - анафаза
, во время которой дочерние хромосомы (хроматиды одной хромосомы) расходятся к разным полюсам клетки.
Следующая стадия деления клетки - телофаза
. Она начинается после того, как дочерние хромосомы, состоящие из одной хроматиды, достигли полюсов клетки. На этой стадии хромосомы вновь деспирализуются и приобретают такой же вид, какой они имели до начала деления клетки в интерфазе (длинные тонкие нити). Вокруг них возникает ядерная оболочка, а в ядре формируется ядрышко, в котором синтезируются рибосомы . В процессе деления цитоплазмы все органоиды (митохондрии , комплекс Гольджи , рибосомы и др.) распределяются между дочерними клетками более или менее равномерно.
Таким образом, в результате митоза из одной клетки получаются две, каждая из которых имеет характерное для данного вида организма число и форму хромосом, а следовательно, постоянное количество ДНК .
Весь процесс митоза занимает в среднем 1-2 ч. Продолжительность его несколько различна для разных видов клеток. Зависит он также от условий внешней среды (температуры, светового режима и других показателей).
Биологическое значение митоза заключается в том, что он обеспечивает постоянство числа хромосом во всех клетках организма. Все соматические клетки образуются в результате митотического деления, что обеспечивает рост организма. В процессе митоза происходит распределение веществ хромосом материнской клетки строго поровну между возникающими из нее двумя дочерними клетками. В результате митоза все клетки организма получают одну и ту же генетическую информацию.
Временной ход митоза и цитокинеза, типичный для клетки млекопитающего. Точные цифры для разных клеток различны. Цитокинез берёт своё начало в анафазе и завершается, как правило,
к окончанию телофазы
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой. М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую. Первые пять профаза, прометафаза, метафаза, анафаза и телофаза составляют митоз, а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза и телофаза. Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза и анафаза. Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.
Профаза

К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.
Прометафаза

Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10-20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона около 30 минут.
Метафаза
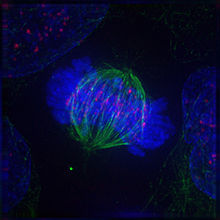
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.
Анафаза

Анафаза самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью достигающей 0,5-2 мкм/мин., при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно в области кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5-2 раза по сравнению с метафазным. В некоторых других клетках анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным. В растительных клетках анафаза В отсутствует.
Телофаза

Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской клетки цитокинезом. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и деление путём образования клеточной пластинки. Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.
| Прево, Жан-Луи |